A Sand Fly, Lutzomyia shannoni Dyar (Insecta: Diptera: Psychodidae: Phlebotomine)
The Featured Creatures collection provides in-depth profiles of insects, nematodes, arachnids and other organisms relevant to Florida. These profiles are intended for the use of interested laypersons with some knowledge of biology as well as academic audiences.
Introduction
Phlebotomine sand flies are of considerable public health importance because of their ability to transmit several viral, bacterial, and protozoal disease-causing organisms of humans and other animals.
Confusion with other types of biting flies is often caused because the common name "sand fly" is also used for other biting flies of genera Ceratopogon and Culicoides. There are about 700 species of phlebotomine sand flies of which about 70 are considered to transmit disease organisms to people (Adler and Theodor 1957).
Sand flies are characterized by their densely hairy wings, giving them a moth-like appearance. Phlebotomines are distinguished from other members of the family by the way they hold their wings erected above the body in a vertical "V", whereas members of other psychodid subfamilies hold their wings flat and near the body (Azar and Nel 2003).
Lutzomyia shannoni Dyar is a proven vector of vesicular stomatitis virus and a suspected vector of visceral leishmaniasis and sand fly fever in Florida. It is one of the more thoroughly studied species of phlebotomine sand flies in North America.
Distribution
Sand flies occur in a wide range of habitats and individual species often have very specific habitat requirements. Lutzomyia shannoni is distributed from Argentina to the United States, including Brazil, Columbia, Panama and Costa Rica. Its distribution is highly disjunct within the range, depending on locally occurring environmental factors such as frequency of precipitation, temperature, physical barriers, habitat availability, and the distribution and abundance of vertebrate hosts (Young and Arias 1992).
In the United States, it has been found throughout the southern states from Florida to Louisiana plus Arkansas, Tennessee, South and North Carolina. This species has been found as far north as Maryland and Delaware. Lutzomyia shannoni is associated with live oak (Quercus viginiana Miller) forests and to a lesser extent, mixed hardwood forests where an abundance of tree holes provides diurnal resting sites for adults. Three other species of Phlebotomine sand flies, L. cubensis (Fairchild and Hertig), L. vexator (Coquillett) and L. cruciata (Coquillett) are also known to occur in Florida (Young and Perkins 1984).
Description
Eggs
The eggs are elongated oval, dark brown in color with polygonal sculpturing over the chorion. The eggs are approximately 0.31 mm long and 0.10 mm wide (Ferro et al. 1998).
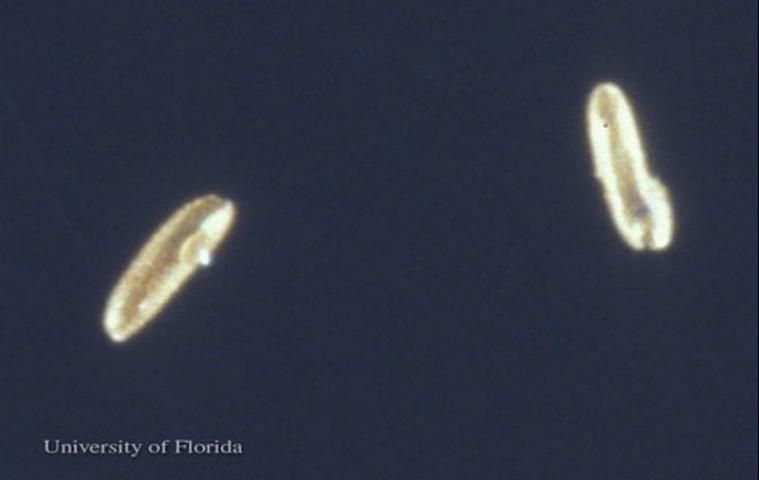
Credit: Jerry Butler, UF/IFAS
First instar larvae
The body color is pale cream, while the head, caudal lobule and caudal setae are light brown. The body length measures between 0.40 and 0.68 mm. The two caudal setae, just after eclosion, measure approximately twice the length of the body. The hypognathous head is well developed and measures between 0.06 and 0.08 mm wide and between 0.04 and 0.05 mm long. The antennal tubercle is larger than the complete antenna. The first antennal segment is long and thin; the second is almost the same size in oval form, with a hair or arista (Ferro et al. 1998).

Credit: Jerry Butler, UF/IFAS
Second instar larvae
The color is similar to that of the first instar but it is larger in size, with four caudal setae approximately twice the body length (Ferro et al. 1998).
Third instar larvae
Third instar larvae are larger than the second instar, but color is similar to that of the first two instars. These larvae have four caudal setae and a Y-form dorsal suture on the head capsule (Ferro et al. 1998).
Fourth instar larvae
The caudal setae are approximately the same as the body length measuring between 2.2 and 2.5 mm. Four well-defined stemmata and two setae with a round and prominent base are present on each side of the head and in the mandibles. The tergal plaque of the eighth abdominal segment and the caudal lobule are black in color. There are no locomotive organs on the thorax (Ferro et al. 1998).
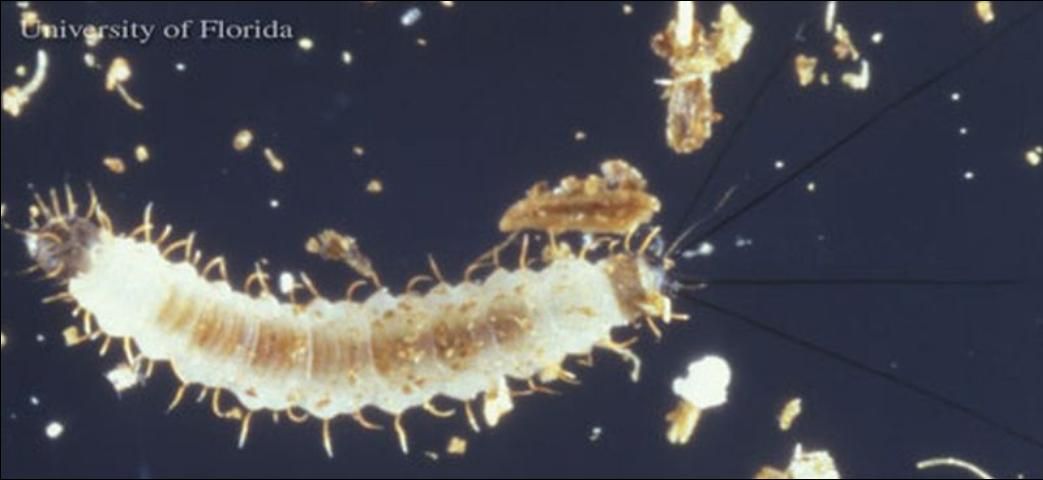
Credit: Jerry Butler, UF/IFAS
Pupae
Pupae are obtect, measuring approximately 2.6 mm in length looking much like a butterfly chrysalis. They are glued to the substrate by the terminal abdominal segments consisting of the terminal pseudopodium of the ninth abdominal segment and the entire exuvium of the fourth instar larva (Ferro et al. 1998).

Credit: Jerry Butler, UF/IFAS
Adults
Adults are small, silvery-brownish, long-legged flies with narrow bodies. The wings are less than 3 mm long, and are held erect above the body. They are nocturnally active insects with weak, direct flight capability, typically not dispersing more than a half kilometer (Morrison et al. 1993). Adults shelter during the day in dark, humid places such as tree holes, animal burrows or under rocks. Sexual dimorphism is marked between the male and female flies. Males have conspicuous external terminalia with a relatively small and slender abdomen compared to the female. Mandibles are absent in males (Young and Perkins 1984).

Credit: Jerry Butler, UF/IFAS

Credit: Jerry Butler, UF/IFAS

Credit: Jerry Butler, UF/IFAS
Life Cycle
Lutzomyia shannoni is an holometabolous insect, progressing from egg to larva, pupa and adult. The complete life cycle can occur in 36 to 74 days depending upon the temperature and larval food quality. The average duration for the egg, larvae (all instars), pupae and adult ranges between 6 to 13, 21 to 51, 7 to 32, and 4 to 15 days respectively (Ferro et al. 1998). The eggs are laid (dispersed singly) in terrestrial microhabitats rich in organic matter that provides food for the larvae. The females require vertebrate blood for the maturation of the follicles. The first oviposition begins between four and eight days after blood ingestion. However, females from Florida are often autogenous (Young and Perkins 1984) and can deposit up to 40 viable eggs without having previous blood meal. Both males and females obtain sugar meals by directly feeding on the tissues of plants and from the honey dew excreted by aphids and coccids. Males do not take a blood meal.
Hosts
The males and females feed on nectar and other plant juices, but females require a blood meal in order to mature a second batch of eggs. The blood meal hosts include white-tailed deer, horses, donkeys, mules, cattle, swine, raccoons, rodents, birds, and humans (Young and Perkins 1984; Comer et al. 1994).
Medical Importance
At least 60 species in the Old World genus Phlebotomus or New World genus Lutzomyia are vectors of several vertebrate pathogens, including a group of parasitic flagellate protozoa, Leishmania spp., which may cause cutaneous, visceral or muco-cutaneous Leishmaniasis; the bacterium, Bartonella bacilliformis causing Oroya fever; and several arboviruses causing sand fly fever and vesicular stomatitis (Lane 1993).
Laboratory studies indicate that at least three species of Leishmania parasites, Leishmania mexicana Biagi, L. panamensis Lainson & Shaw, and L. chagasi Cunha & Muniz, can develop in Lutzomyia shannoni (Ferro et al. 1998). Lutzomyia shannoni feeds on mammals, including humans, and has been reported to transmit a visceral leishmaniasis in dogs, hamsters, and other mammals. This pathogen can also be transmitted to humans through contact with the organism, though human infection acquired as a result of direct exposure to an infected dog has never been reported (Lawyer and Young 1987; Travi et al. 2002; Schantz 2007).
Visceral leishmaniasis had not been considered established in North America, however, recent events indicated that it does occur in foxhounds and perhaps certain other breeds of dogs. These foxhounds appear to have been infected with a strain of the parasite that originated in southern Europe. The Spinone dog breed is at risk of the infection because many dogs of this breed have been directly imported from Italy where the infection is present. Transmission through L. shannoni has not been confirmed in North America and it is suspected that the infection is transmitted from dog to dog in blood and secretions incidental to biting, licking, breeding, blood transfusion, etc. (Duprey et al. 2006; Schantz 2007). The signs of leishmaniasis in dogs are variable and often do not appear until many months after exposure to the infection. The infected dog may lose weight and develop swollen lymph nodes or anemia. Skin lesions are common including hair loss and ulcers. The dog may lose appetite, become lethargic and bleed from the nose. Medical treatments can improve condition and prolong survival, but rarely eradicate the infection. Without treatment most infected dogs ultimately die (Schantz 2007). In humans, signs and symptoms include fever, weight loss, anaemia and substantial swelling of the liver and spleen (WHO 2015).
Lutzomyia shannoni is a proven vector of New Jersey serotype of vesicular stomatitis virus (VSV) on Ossabaw island, Georgia, US (Comer et al. 1994; Stallknecht 2000). Vesicular stomatitis is an acute viral vesicular disease of cattle, horses, deer and pigs (and occasionally humans) causing fever 24 to 48 hours after infection, vesicular lesions on mouth, coronary bands, and teats. Rupture of the oral vesicles leads to pain, excessive salivation and inappetance. The coronary band lesions lead to lameness. VSV infection resembles foot-and-mouth disease when it occurs in cattle and the other viral vesicular diseases when it occurs in pigs. VSV causes flu-like symptoms less commonly, oral vesicles and cervical lymphadenopathy in humans. Humans can become infected with VSV when handling infected animals if proper biosafety procedures are not followed. Prevalence of this disease in humans may be under-reported because it may often go undetected or may be misdiagnosed (Nebraska Department of Agriculture 2007).
Management
Measures to control adult sand flies (Lutzomyia and Phlebotomus) include the use of insecticides (currently mostly pyrethroids) for residual spraying of dwellings and animal shelters, space-spraying, insecticide-treated nets, impregnated dog-collars and personal protection through application of repellents/insecticides to skin or fabrics.
Spraying of residual insecticides on surfaces in the home has been the primary method for control of sand flies but is obviously ineffective for those species which bite away from residences. This control technique has also been used for killing Anopheles mosquitoes that transmit malaria and in some regions is effective in reducing both malaria and leishmaniasis (Alexander and Maroli 2003).
Certain species of mammals are important reservoirs of Leishmania and by eliminating the reservoir species living near human habitation, disease rates can sometimes be decreased. For instance, rodenticides have been used against the great gerbil, Rhombomys opimus, in Central Asia. Insecticide spraying of larval habitat is usually normally not possible because very little is known about where the larvae occur (Ashford 1999).
Selected References
Adler S, Theodor O. 1957. Transmission of disease agents by phlebotomine sand flies. Annual Review of Entomology 2: 203–226.
Alexander B, Maroli M. 2003. Control of phlebotomine sandflies. Medical and Veterinary Entomology 17: 1–18.
Ashford RW. 1999. Cutaneous leishmaniasis: strategies for prevention. Clinics in Dermatology 3: 327–332.
Azar D, Nel A. 2003. Fossil psychodoid flies and their relation to parasitic diseases. Memórias do Instituto Oswaldo Cruz 98: 35–37.
Comer JA, Irby WS, Kavanaugh DM. 1994. Hosts of Lutzomyia shannoni (Diptera: Psychodidae) in relation to vesicular stomatitis virus on Ossabaw Island, Georgia, U.S.A. Medical and Veterinary Entomology 8: 325–30.
Duprey ZH, Steurer, FJ, Rooney JA, Kirchhoff LV, Jackson JE, Rowton ED, Schantz PM. 2006. Canine visceral leishmaniasis, United States and Canada, 2000-2003 (RESEARCH). Emerging Infectious Diseases 12: 440–446.
Fasulo TR. (2002). Bloodsucking Insects and Filth-breeding Flies. Bug Tutorials. University of Florida/IFAS. CD-ROM. SW 156.
Ferro C, Cardenas E, Corredor D, Morales A, Munstermann LE. 1998. Life cycle and fecundity analysis of Lutzomyia shannoni (Dyar) (Diptera: Psychodidae). Memórias do Instituto Oswaldo Cruz 93: 195–199.
Lane RP. 1993. Sand flies (Phlebotominae). In Medical Insects and Arachnids (eds. R.P. Lane RP, and Crosskey RW). Chapman and Hall, London, pp. 78–119.
Lawyer PG, Young DG. 1987. Experimental transmission of Leishmania mexicana to hamsters by bites of phlebotomine sand flies (Diptera: Psychodidae) from the United States. Journal of Medical Entomology 24: 458–462.
Morrison AC, Ferro C, Morales A, Tesh RB, Wilson M L. 1993. Dispersal of the sand fly Lutzomyia longipalpis (Diptera: Psychodidae) at an endemic focus of visceral leishmaniasis in Colombia. Journal of Medical Entomology 30: 427–435
Nebraska Department of Agriculture. (2011). Vesicular Stomatitis. Animal Industry. https://nda.nebraska.gov/animal/diseases/vs/index.html.
Schantz PM. 2004. Visceral leishmaniasis in dogs. Spinone Club of America. http://www.spinone.com/Health/Leishmaniasis/cdc_Schantz.htm (17 October 2007).
Stallknecht DE. 2000. VSV-NJ on Ossabaw Island, Georgia: The truth is out there. Annals of the New York Academy of Sciences 916: 431–436.
Travi BL, Ferro C, Cadena H, Montoya-Lerma J, Adler GH. 2002. Canine visceral leishmaniasis: dog infectivity to sand flies from non-endemic areas. Research in Veterinary Science 72: 83–6.
Young DG, Arias JA. 1992. Flebótomos: vectores de leishmaniasis en las Américas, Org. Pan-Amer, Salud, Tecnical Report No. 33
Young DG, Perkins PV. 1984. Phlebotomine sand flies of North America (Diptera: Psychodidae). Mosquito News 44: 263–306.
World Health Organization (WHO). (2015). Leishmaniasis. https://www.who.int/health-topics/leishmaniasis (27 September 2022).

