Velvet Ants, Mutillidae (Insecta: Hymenoptera)
The Featured Creatures collection provides in-depth profiles of insects, nematodes, arachnids and other organisms relevant to Florida. These profiles are intended for the use of interested laypersons with some knowledge of biology as well as academic audiences.
Introduction
Insects in the family Mutillidae are often referred to as "velvet ants" because female members of the family lack wings and have coarse setae that cover most of their body, making them resemble hairy ants. To the surprise of most people, mutillids are not ants at all—they are wasps.

Credit: Lyle Buss, UF/IFAS

Credit: James Castner, UF/IFAS

Credit: Lyle Buss, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Clemson University, www.forestryimages.org
Distribution
The Mutillidae family contains approximately 230 genera/subgenera and about 8,000 species worldwide (Manley and Pitts 2002). Approximately 435 species occur in mostly arid areas of the southern and western parts of North America (Triplehorn and Johnson 2005). Fifty species in seven genera are found in Florida (Table 1) (Krombein 1979).
Biology
All Mutillidae are solitary parasitoid wasps that mostly attack mature larvae or pupae of other solitary Hymenoptera. However, velvet ants have been observed targeting non-feeding stages of Diptera, Coleoptera, Lepidoptera, Blattodea, and even some eusocial Hymenoptera (Brothers et al. 2000).
Finding a matching reproductive pair can be difficult, if not impossible for some species, because of the extreme sexual dimorphism displayed within this family. Color patterns and the relative body size of the two sexes of the same species can be very different, making it very difficult to relate one sex with the other (Mickel 1928).
In Florida, male Mutillidae are frequently larger (heavier) than the females. One study in south-central Florida found that in seven out of 13 species, the males were heavier than the females. The remaining six species either showed comparable sizes between the male and female (3) or the female was larger than the male (3) (Deyrup and Manley 1986).
Biological data is very scarce and differences obviously exist between genera and species, especially with regards to their life cycles and development, but the following has been reported for most Mutillidae. Females have the difficult job of locating potential hosts. Suitable hosts may be difficult to find for several reasons including: concealment, low population densities when solitary and lastly, if the hosts are eusocial, they will be heavily defended. Once a host is located, female velvet ants parasitize the "hard" life stages (i.e., hardened pre-pupae, pupae, ootheca, eusocial cells, and cocoons) of the hosts and the emerging velvet ant larvae are essentially ectoparasites of those life stages. (Brothers et al. 2000). Once development is completed, adults leave the nest and seek a mate.
Identification
All female Mutillidae are wingless and resemble ants. They can easily be differentiated from ants by the lack of petiole nodes, which are present on all ant species. In addition, the mesosomatic (thoracic) segments are completely fused and have at most two segments. The metasoma (abdomen) contains six visible terga (dorsal surface of any body segment) and a "felt line" of dense, closely appressed hairs is located laterally on the second metasomatic tergum (except for the genus Ephuta) (Manley and Pitts 2002, Triplehorn and Johnson 2005).
Unlike the female, the male meso- and meta-thorax are not fused and they have three mesosomatic segments. Their metasomal region contains seven visible terga and a felt line is also on the lateral margin of tergite II. All male Mutillidae found in Florida normally have large wings lacking jugal lobes and forewings with the M and Cu1 veins ending far from the apical margin (Manley and Pitts 2002).
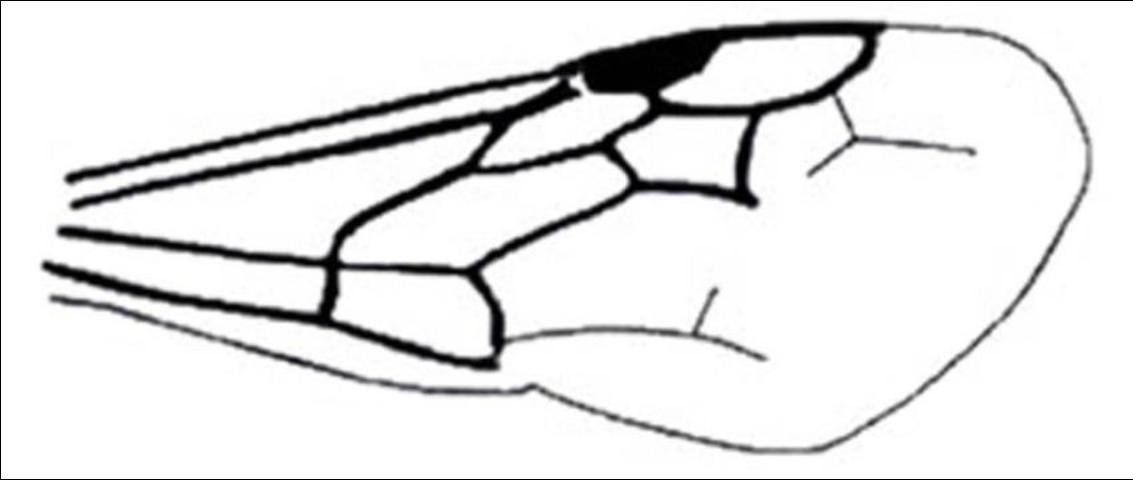
Credit: Manley and Pitts 2002
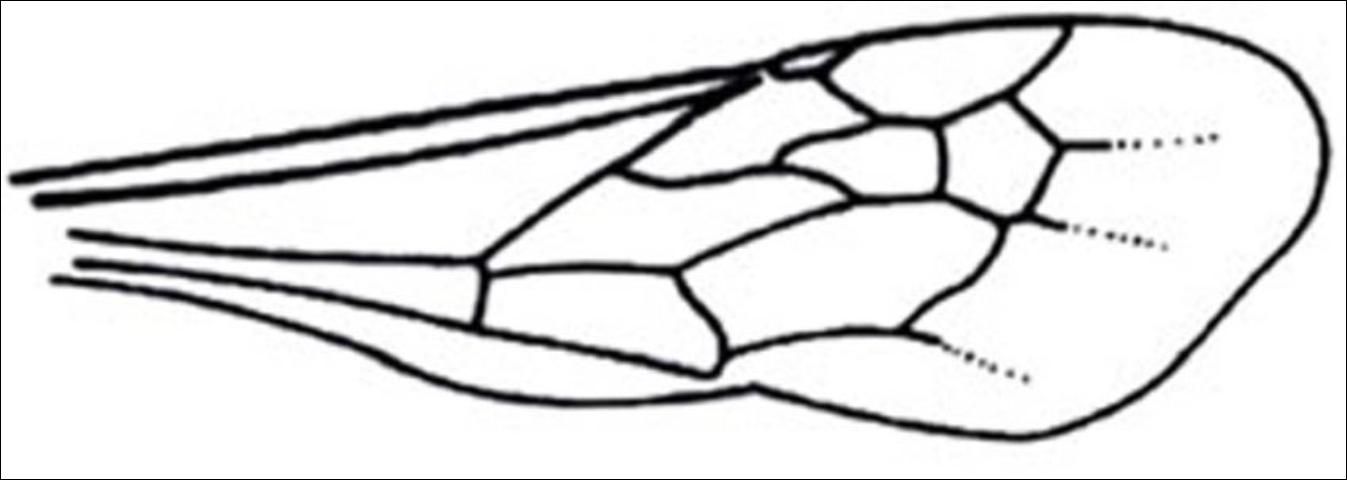
Credit: Manley and Pitts 2002
Key to Florida Genera
The most recently published key to genera and subgenera was issued by Manley and Pitts (Manley and Pitts 2002). Conclusions and pictures contained within this key were derived directly from this publication, but only for genera found in Florida.
Female Specimens
1. Metasomal segment I completely sessile with second (without petiole) . . . . . 2
1'. Metasoma petiolate or at most subsessile, with definite constriction between first two segments (Figure 9) . . . . . 5
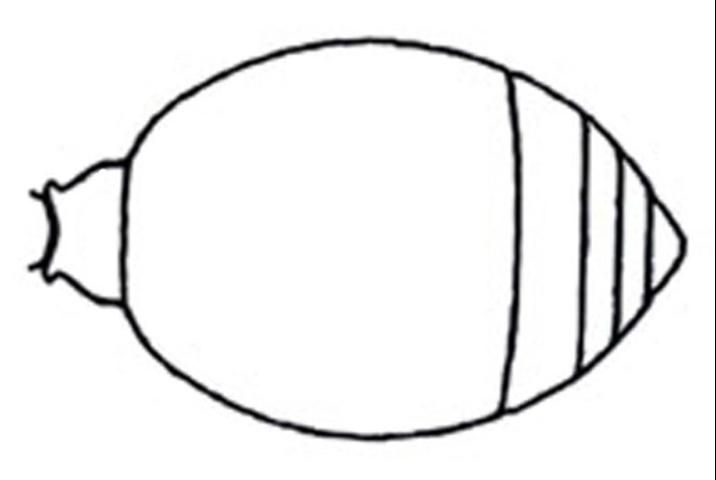
Credit: Manley and Pitts 2002
2. Eyes strongly ovate; mesosoma long, rectangular, generally narrowed medially (Figure 10); tergite II generally maculated with two spots or lines of pale setae . . . . . Timulla spp.
2'. Eyes circular to slightly ovate; mesosoma otherwise (short, rectangular or narrowed posteriorly, if narrowed medially, mesosoma pyriform not rectangular); tergite II not maculated with spots of pale setae . . . . . 3
Credit: Manley and Pitts 2002
3. Head, in dorsal view, less than twice the width of mesosoma (Figure 11); pygidium usually with lateral carinae, mesosoma narrowed medially; head not dentiform (toothed); if hypostomal teeth present they are not prominent, nor bent apically; posterolateral angles of head, usually not strongly carinate (ridged) . . . . . Pseudomethoca
3'. Mesosoma as wide or wider than head in dorsal view, rectangular without fine carina between pronotum and mesonotum (Figure 12); eyes slightly ovate; mandible ventrally toothed or not; sternal felt line absent; plumose setae present or absent; integument visible, usually sparsely covered with appressed and erect simple setae; pygidium variable . . . . . 4
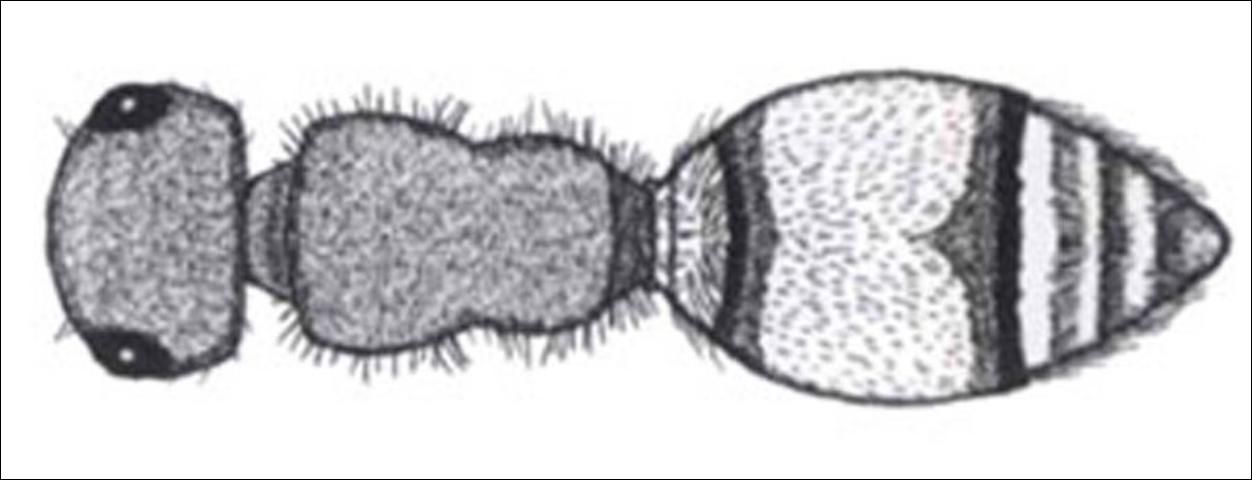
Credit: Manley and Pitts 2002

Credit: Manley and Pitts 2002
4. Pygidium dull, shagreened (rough), with parallel carinae only on basal two-thirds or less . . . . . Photomorphus, subgenus Photomorphina
4'. Pygidium smooth and shiny, with complete parallel carinae on disk . . . . . Photomorphus, subgenus Photomorphus
5. Felt lines absent on metasomal tergite II; petiole short, transverse, parallel-sided (Figure 13); eyes distinctly ovate; a band of silvery, dense, pubescence present at apex of petiole and metasomal segment II; small; densely punctuate (subfamily Mutillinae, tribe Ephutini) . . . . . Ephuta
5'. Felt lines present on metasomal tergite II; petiole not transverse or parallel sided; eyes circular to slightly ovate; other characters variable . . . . . 6
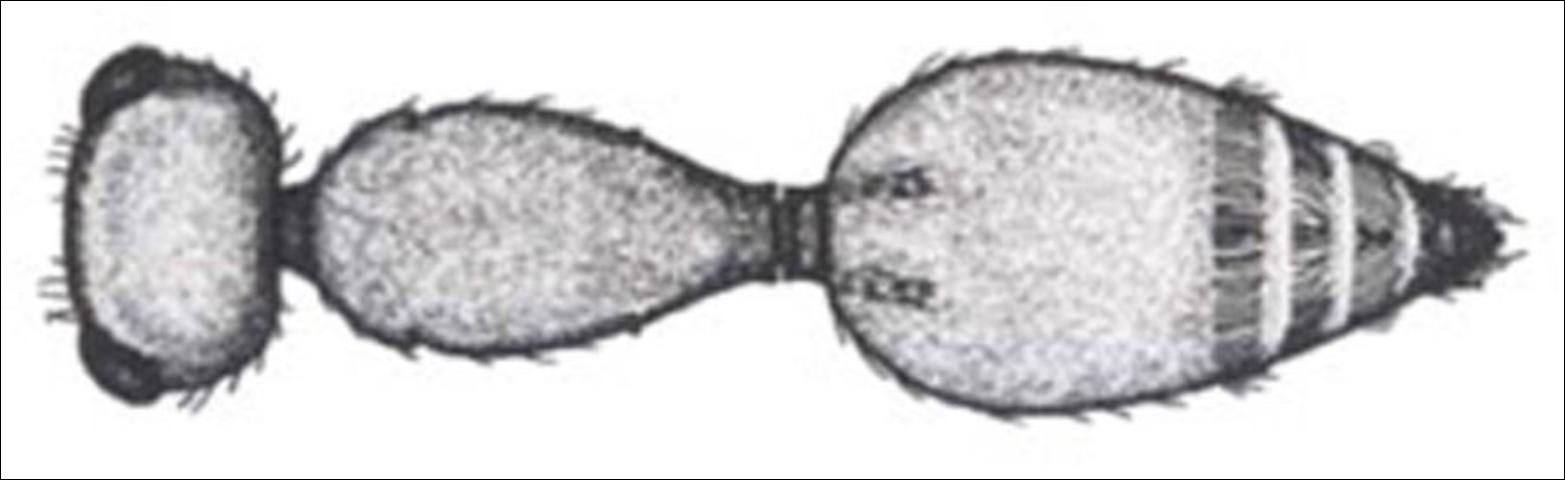
Credit: Manley and Pitts 2002
6. Plumose setae limited to area of short dense white setae on dorsum of petiole, and apical fringe of tergite II (Figure 14); pygidial area undefined laterally by carinae; genal carina absent; flagellomere II greater than 1.75x length of first; antennal scrobe distinctly carinate dorsally; propodeum elongate . . . . . Sphaeropthalma, subgenus Sphaeropthalma
6'. Plumose setae totally absent . . . . . 7
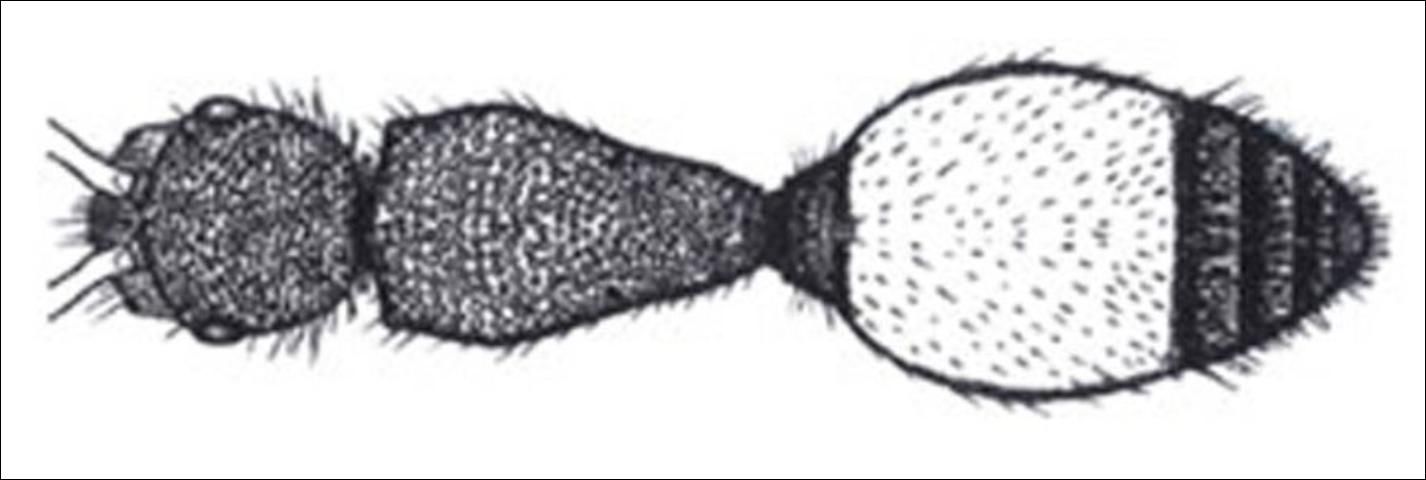
Credit: Manley and Pitts 2002
7. Pygidial area well-defined; petiole not diciform (rounded) (Figure 15) . . . . . Dasymutilla
7'. Pygidial area obsolete, not defined laterally; petiole distinctly disciform (Figure 16); Anterior and propodeal spiracles tuberculate (Figure 17) . . . . . Lomachaeta

Credit: Manley and Pitts 2002
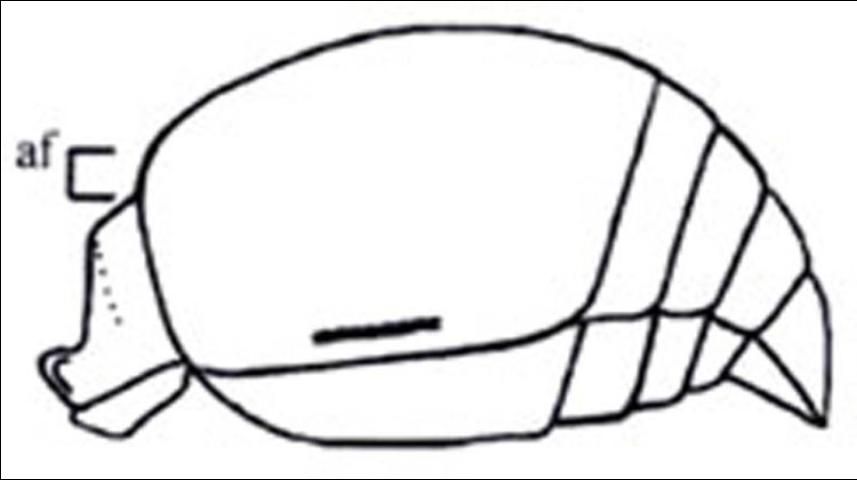
Credit: Manley and Pitts 2002
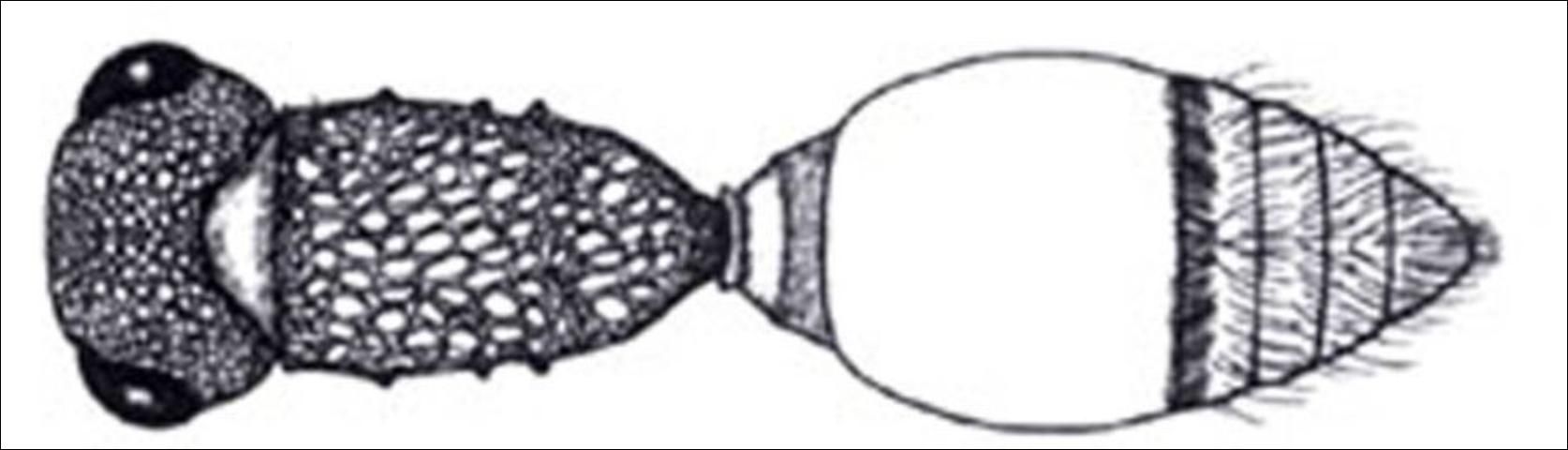
Credit: Manley and Pitts 2002
Male Specimens
1. Compound eye approximately round, not emarginated, smooth and shiny (Figure 18); pterostigma sclerotized . . . . . 2
1'. Compound eye with inner margin deeply and sharply emarginated (Figures 19, 20); pterostigma membranous or absent (Figure 21) . . . . . 6
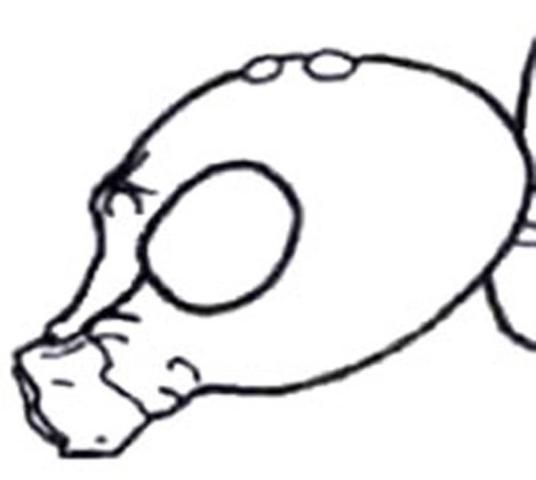
Credit: Manley and Pitts 2002
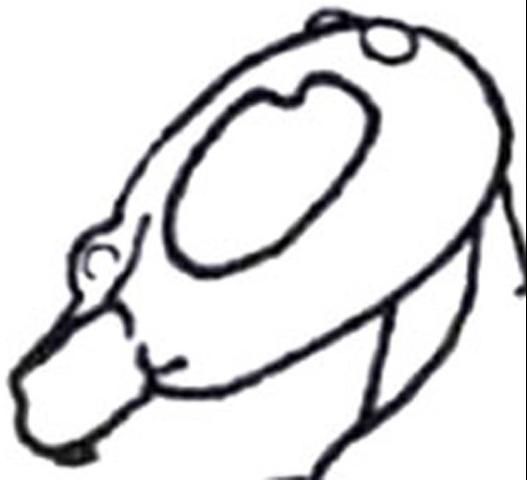
Credit: Manley and Pitts 2002

Credit: Manley and Pitts 2002
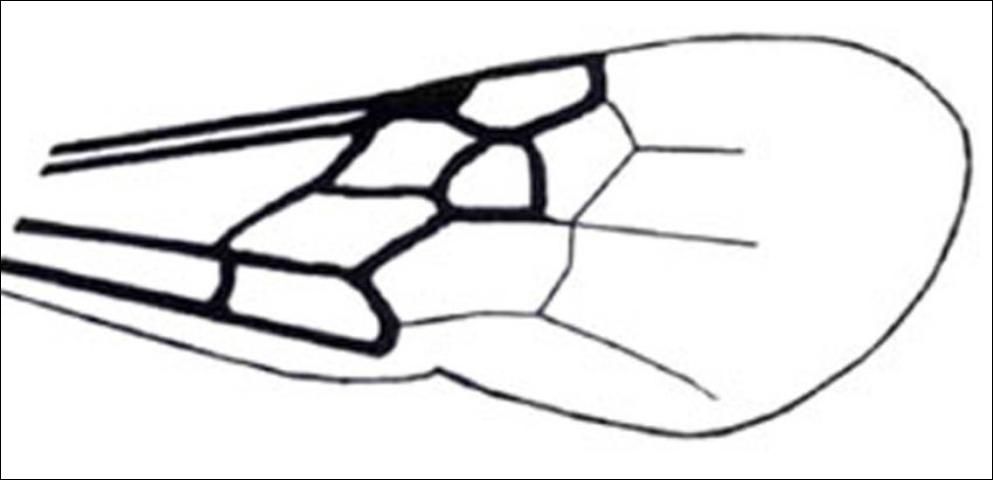
Credit: Manley and Pitts 2002
2. Metasomal segment I completely sessile with second (Figure 22); Felt line present on lateral margin of tergite II only; hypostomal tooth not developed; head less than twice width of mesosoma; posterolateral angles of head not strongly carinate or dentiform; wings macropterous . . . . . Pseudomethoca
2'. Metasoma petiolate or at most subsessile, with definite constriction between first two segments (Figure 23) . . . . . 3
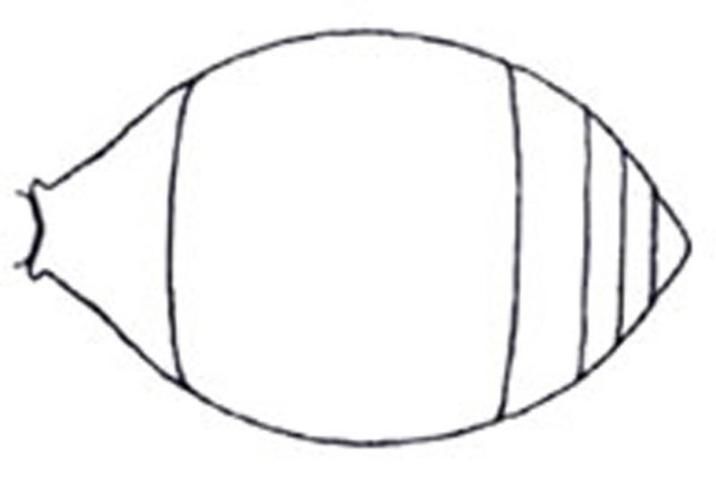
Credit: Manley and Pitts 2002
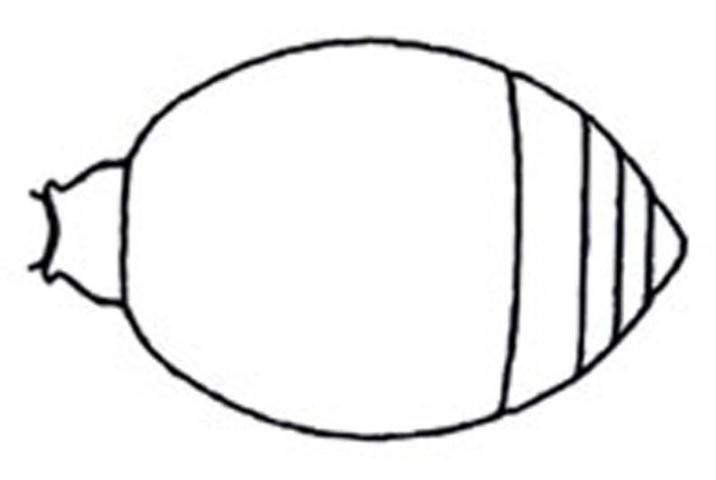
Credit: Manley and Pitts 2002
3. Felt line present on lateral margin of tergite II and sternite II; ventral mandibular tooth present (Figure 24); mesoscutal notauli present (separates mesoscutum into medial and lateral lobes); flagellomere I longer than wide, not like pedicel; hypopygium (last visible abdominal sterum) normal, rounded, lateral margins not carinate or dentiform; mesosternum with pair of longitudinal to transverse dentate ridges anterior to mesocoxae, closer to procoxae; plumose hairs vestigial or absent . . . . . . 6
3'. Felt line present on lateral margin of tergite II only; Mesotibia with two spurs; mesotibia cylindrical, not flattened and arcuate; plumose setae present or absent; mesosternum unmodified . . . . . 4

Credit: Manley and Pitts 2002
4. Mandible tridentate apically, broadly emarginated ventrally with small, distinct tooth; eyes and ocelli small, not protuberant, compound eye distinctly separated from mandible, distance from compound eye to posterolateral angle of head distinctly greater than greatest diameter of eye; notauli present, complete; tergites II-V without row of lanceolate (lance-shaped) bristles at distal margin; pterostigma of forewing conspicuous; marginal cell length equal to or slightly longer than stigma; pygidium and hypopygium short, transverse . . . . . Sphaeropthalma, subgenus Sphaeropthalma
4'. Mandible bidentate or tridentate apically, but not emarginate or toothed ventrally . . . . . 5
5. Notauli absent; subplumose setae present; tergites II-V with row of lanceolate bristles at distal margin; pterostigma of forewing vestigial, inconspicuous (Figure 25) . . . . . Lomachaeta
5'. Tergites II-V without row of lanceolate bristles at distal margin; subplumose setae absent; wing venation normal, not greatly reduced; pterostigma of forewing conspicuous (Figure 21) . . . . . Dasymutilla
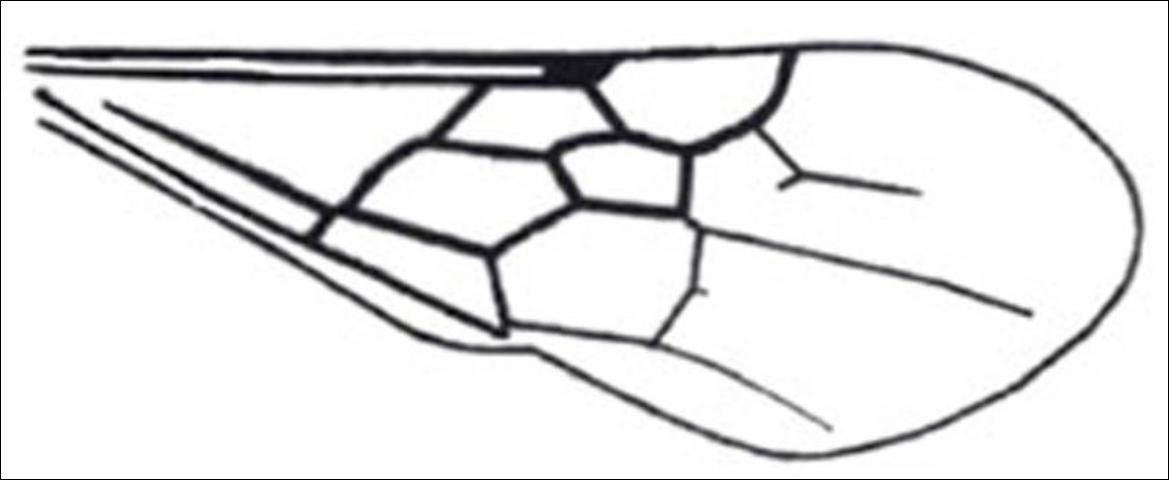
Credit: Manley and Pitts 2002
6. Metasomal segment I sessile with second (Figure 26); humeral angles rounded . . . . . Timulla
6'. Metasomal segment I slender, short, parallel-sided, not sessile (Figure 27); humeral angles angulate, sharply produced; small; densely punctuate . . . . . Ephuta
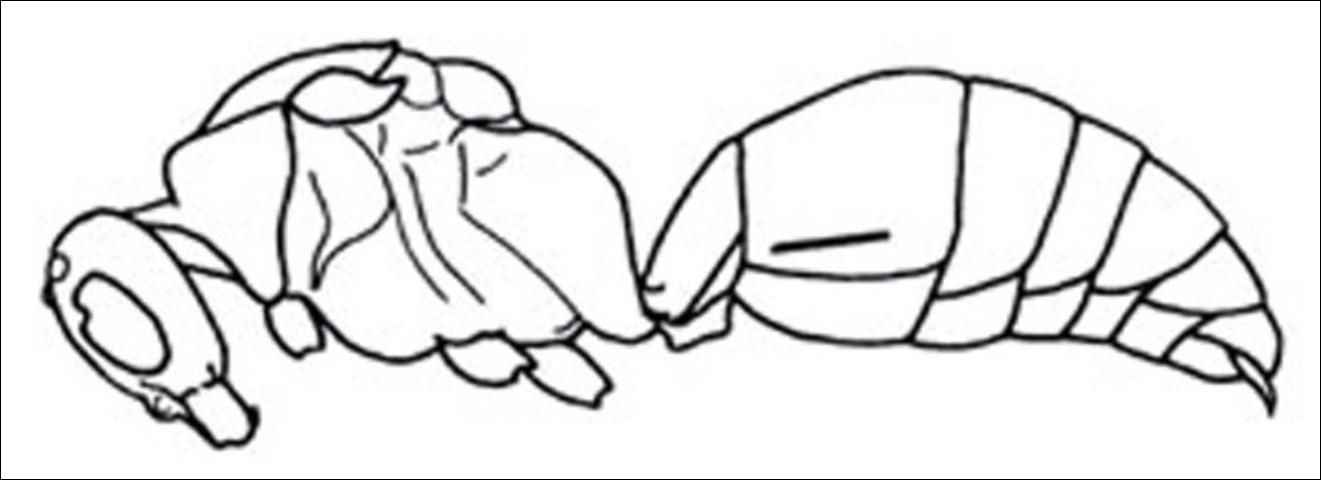
Credit: Manley and Pitts 2002
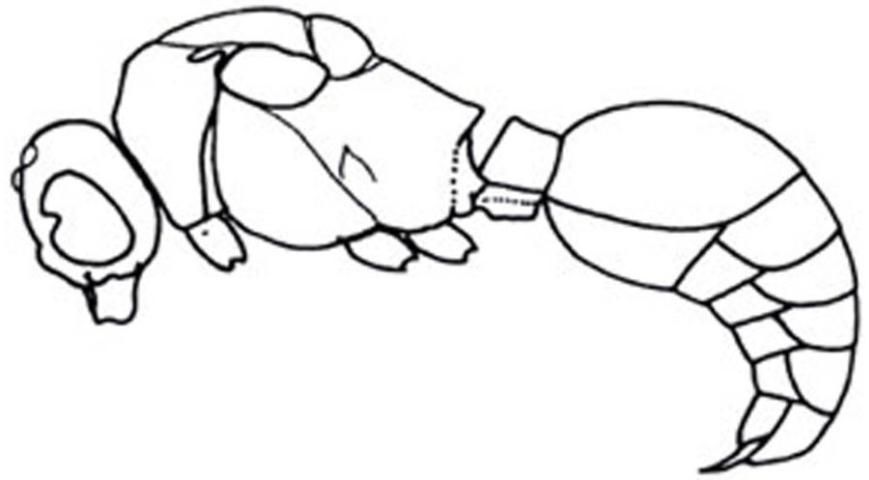
Credit: Manley and Pitts 2002
7. Mesocoxae approximate; mandible tridentate apically; mentum never produced into a distinct process . . . . . Photomorphus, subgenus Photomorphina
7'. Mesocoxae separated; mandible bidentate apically (Figure 24); mentum distinctly produced as an anterior tubercle (small nodule) or posterior lingulate (tongue-shaped) process . . . . . Photomorphus, subgenus Photomorphus
Florida Species
See Table 1

Credit: Lyle Buss, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Sean McCann, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Lyle Buss, University of Florida

Credit: Ken Gray, Oregon State University

Credit: Ken Gray, Oregon State University

Credit: Ken Gray, Oregon State University

Credit: Lyle Buss, University of Florida

Credit: Charles Schurch Lewallen
Pest Status and Management
Mutillidae pose no real threat to humans, livestock, or domesticated animals other than the unfortunate accidental sting associated with unaware victims. Like other stinging insects, only female Mutillidae are capable of stinging. Some species are referred to as "cow killers" or "mule killers," because their sting is said to be painful enough to kill an animal. Despite this assertion, they are not aggressive nor is their venom comparatively toxic. Most female mutillids are so docile, they produce a startle response (an audible stridilation) when provoked to warn potential victims of her sting. The LD50 of their venom when exposed to mice is around 71 mg/kg body weight which is relatively non-toxic compared to the very toxic harvester ant, Pogonomyrmex maricopa, venom (LD50 = 0.12 mg/kg) and the moderately toxic honey bee, Apis mellifera, venom (LD50 = 2.8) (Meyer 1996). No control measures are necessary.

Credit: Jeffrey C. Hertz, University of Florida

Credit: Jeffrey C. Hertz, University of Florida
Acknowledgments
I sincerely thank Dr. Don Manley, of Clemson University, for providing me the most current reference literature and taking the time to review this publication. I also thank Dr. Gavin Broad for graciously giving me permission to use numerous images previously published in the Journal of Hymenoptera Research.
Selected References
Brothers DJ, Tschuch G, Burger F. 2000. "Associations of mutillidae wasps (Hymenoptera, Mutillidae) with eusocial insects." Insectes Sociaux 47: 201–211.
Deyrup MA, Manley DG. 1986. "Sex-biased size variation in velvet ants (Hymenoptera: Mutillidae)." Florida Entomologist 69: 327–335.
Krombein KV, et. al. 1979. Catalog of Hymenoptera in America north of Mexico. Smithsonian Institution Press, Washington.
Manley DG, Deyrup MA. 1987a. "A new species of Dasymutilla (Hymenoptera: Mutillidae) from Florida." Journal of Entomological Science 22: 123–125.
Manley DG, Deyrup MA. 1987b. "A new species of Photomorphus (Hymenoptera: Mutillidae) from Florida." Journal of Entomological Science 22: 57–60.
Manley DG, Pitts JP. 2002. "A key to genera and subgenera of Mutillidae (Hymenoptera) in America north of Mexico with description of a new genus." Journal of Hymenoptera Research 11: 72–100.
Meyer WL. 1996. Most toxic insect venom. University of Florida Book of Insect Records. https://entnemdept.ufl.edu/walker/ufbir/chapters/chapter_23.shtml (28 February 2022)
Mickel CE. 1928. "Biological and taxonomic investigations on the mutillid wasps." Bulletin of the U.S. National Museum 143: 351.
Pitts JP, Manley DG. 2004. "A revision of Lomachaeta Mickel, with a new species of Smicromutilla Mickel (Hymenoptera: Mutillidae)." Zootaxa 474: 1–27.
Triplehorn CA, Johnson NF. 2005. Family Mutillidae. In Borror and Delong's Introduction to the Study of Insects, 7th ed. Thomson Brooks/Cole, Belmont, CA.

