Classical Biological Control of Brazilian Peppertree (Schinus terebinthifolia) in Florida
Brazilian peppertree, Schinus terebinthifolia Raddi (Anacardiaceae), also known as Christmasberry, Florida Holly, and aroeira (Brazil), is an aggressive, rapidly colonizing invasive weed of disturbed habitats, natural communities and conservation areas in peninsular Florida (Cuda et al. 2004, Cuda et al. 2006, Manrique et al. 2013). Native to Argentina, Paraguay, and Brazil, Brazilian peppertree was introduced into Florida as a landscape ornamental in the late 19th century. The popularity of Brazilian peppertree as an ornamental plant can be attributed to the numerous bright red drupes (fruits) produced during the October to December holiday season (Figure 1). Grown as a substitute for the more traditional English holly (Ilex aquifolium L.), Brazilian peppertree was common in cultivation in Florida during the first half of the 20th century. However, this relative of poison ivy was a rare component of the native flora in Florida until the late 1950s when the first naturalized plants were discovered in Monroe County.

Credit: A. Murray, University of Florida, Center for Aquatic and Invasive Plants (used with permission).
Brazilian peppertree currently dominates entire ecosystems in south-central Florida (Cuda et al. 2006). It is considered an important invader of the Everglades National Park and poses a significant threat to ongoing Everglades restoration efforts (Rodgers et al. 2014). Brazilian peppertree is also one of the costliest upland invasive plants in Florida. State agencies spend nearly $3 million each year for its control (Hiatt et al. 2019). Once established, Brazilian peppertree quickly displaces the native vegetation, often forming dense monocultures that reduce the biological diversity of plants and animals in the invaded area. Although herbicides (https://edis.ifas.ufl.edu/publication/aa219) and mechanical or physical control practices (e.g., cutting, burning, and flooding) are routinely used often in combination for controlling existing stands (https://bugwoodcloud.org/CDN/fleppc/publications/BrazillianPeppertree_Final_Oct19.pdf), these conventional methods are expensive, labor intensive, and provide only temporary control due to the plant's regenerative capacity. Furthermore, nonselective chemical and mechanical controls are unsuitable for sensitive natural areas (e.g., coastal mangrove forests, The Everglades) because they can have negative effects on nontarget species and increase water pollution. Minimizing the use of herbicides and other nonselective control practices is needed to maintain the integrity of Florida's fragile environment and natural resources. Biological control—the introduction of host-specific natural enemies into Florida that are capable of selectively damaging Brazilian peppertree—will accomplish this goal.
In the 1980s, Brazilian peppertree was identified as a suitable target for classical biological control (Habeck et al. 1994). Biological control is an appropriate management tactic because the invasive characteristics exhibited by Brazilian peppertree are consistent with the enemy release hypothesis (Williams 1954). The key elements of this hypothesis are that (a) native, host-specific enemies control the abundance and distribution of native plants; (b) escape from host-specific enemies is a key contributor to invasive plant success; and (c) enemy escape benefits invasive plants because they gain a competitive advantage over native plants as a result of being liberated from their herbivores. Also, because no native species in the genus Schinus occur in the United States, the potential for nontarget damage by approved biological control agents would be low.
Several insects have been identified from exploratory surveys conducted in Argentina, Brazil, and Paraguay as potential biological control agents of Brazilian peppertree (Habeck et al. 1994; Hight et al. 2002; Cuda et al. 2006; McKay et al. 2009; Wheeler et al. 2016). The following insects were selected for further study because they visibly damage the plant in its native range and were collected only from Brazilian peppertree or a few closely related species during field surveys.
Brazilian Peppertree Thrips, Pseudophilothrips ichini
The biology and field host range of Pseudophilothrips ichini (Hood) (Thysanoptera: Phlaeothripidae) were studied in southeastern Brazil (Garcia 1977). Pseudophilothrips ichini has not been observed feeding on plants other than Brazilian peppertree in its native range (Garcia 1977, J. H. Pedrosa, personal observation). Because this thrips was found attacking only Brazilian peppertree in field surveys, Garcia (1977) suggested that P. ichini might be a good candidate for biological control of this invasive weed. The original laboratory colonies of P. ichini were found to be a complex of two cryptic species (Cuda et al. 2009, Mound et al. 2010, Cuda 2016). After many investigations into the species complex and determination of which of the species was most adapted to the populations of Brazilian peppertree in Florida, it was determined that Pseudophilothrips ichini was the best fit for use as a biological control agent. The other species in the cryptic species complex was later identified as Pseudophilothrips gandolfoi Mound (Mound et al. 2010). This species had very low survival rates on Brazilian peppertree plants in Florida (Manrique et al. 2008).
The life cycle of P. ichini begins when the female deposits her eggs on the young leaves and stems of Brazilian peppertree. After hatching, the immature thrips undergo two larval instars that are active feeding stages. The wingless larvae are orange or yellow in color (Figure 2). As soon as the larval feeding phase is completed on the host plant, the remainder of the life cycle occurs in or on the soil. Unlike other families of the Thysanoptera that have only two pupal instars (the propupa and pupa), thrips belonging to the family Phlaeothripidae that includes P. ichini are unique in that they undergo three nonfeeding pupal instars (the propupa, pupa I and pupa II) instead of two (Mound and Marullo 1996). Although these developmental phases are not true larvae or pupae, these terms are commonly used to describe the immature stages in a thrips life cycle.

Credit: (adult) M. Vitorino, University of Blumenau; (larvae) V. Manrique, University of Florida
Adults of P. ichini are black, winged, and relatively small (3–6 mm) (Figure 2) but have a high reproductive rate. Pseudophilothrips ichini is polyvoltine; up to four generations per year have been observed in Curitiba, Brazil, and it is considered a common species in its native range (Garcia 1977). Mating is not required to produce offspring. Unmated females deposit eggs that develop only into males whereas mated females produce eggs that develop into females (Mound and Marullo 1996). This form of parthenogenetic reproduction is called arrhenotoky.
In Brazil, the adults overwinter on Brazilian peppertree. In early spring (September), females start laying eggs singly or in small groups on the leaflet pedicels and blades, or on the new tender shoot growth. The duration of the immature stages is variable, depending on climate and other factors. The larvae hatch from the eggs in 7–8 days at 24°C (75.2°F). The first and second instars last 6 days and 11–12 days, respectively. The two nonfeeding prepupal and pupal stages require ~ 8 days to complete their development. After transformation to the adult stage, females undergo a 5 to 15 day preoviposition period, and can oviposit up to 220 eggs during their lifetime (45–78 days). Duration of the complete life cycle for P. ichini is temperature dependent. For example, the life cycle from egg to egg was completed in 76 days at 18°C, 38 days at 24°C (75.2°F) (Garcia 1977), and 20 days at 27°C (80.6°F) (Wheeler et al. 2016), Under laboratory conditions, adults lived on average 78 days at 23.1°C (73.4°F) and 50 days at 27°C (80.6°F) (Garcia 1977; Wheeler et al. 2016) when maintained in vials provided with food.
Both the larval and adult stages damage the plant. Larvae of P. ichini usually are found clustered around the stem of a tender shoot (Figure 2). They feed by rasping and sucking the plant sap, which frequently kills the growing tip. Adults are usually found on the new unfolding leaves where they feed, mate, and oviposit. Although they can be more randomly distributed on the plant, adults usually are found aggregated with the developing larvae. Adults also will feed on the flowers, causing them to abort. This type of feeding damage can inhibit seed production in mature plants and has been shown to reduce the growth rate of younger plants (Furmann et al. 2005). In addition, there is anecdotal evidence suggesting that feeding damage by P. ichini promotes infection by plant pathogens that contributes to shoot death (R. Barreto, pers. comm.).
After receiving Brazilian export permits (permit # 07BR001027/DF; 09BR003084/DF; 09BR003939/DF; 10BR004435/DF; 11BR005954/DF; 12BR007880/DF; 14BR013387/DF; 14BR012781/DF; 15BR017495/DF; 19BR030432/DF) and USDA-APHIS-PPQ import permits, the laboratory host range of P. ichini was investigated in approved Florida containment laboratories (Cuda et al. 2009, Wheeler et al. 2017). A petition to release P. ichini from quarantine was initially prepared and submitted to the federal interagency Technical Advisory Group for the Introduction of Biological Weed Control Agents, or TAG (https://edis.ifas.ufl.edu/publication/in607) in November 1996. Request for field release was initially denied because the biological and host range testing data presented in the original petition did not adequately address the risk to native Caribbean plant species and to the closely related California peppertree, Schinus molle L., a common introduced ornamental in southern California. A new petition to release the thrips in Florida was prepared and resubmitted to the TAG in October 2002 (Cuda et al. 2002). The revised petition adequately addressed all of the concerns raised by reviewers in the earlier petition, and the TAG recommended field release of P. ichini in May 2007. However, because of the cryptic species issue (Mound et al. 2010), TAG withdrew the recommendation for field release until the host range of the species complex was resolved. The release of P. ichini from quarantine was finally recommended by TAG in May 2016, and it was released in Florida on 16 July 2019. This is the first insect released for biological control of Brazilian peppertree in the continental United States.
Scientists at UF-IFAS, USDA-Agricultural Research Service, and the Florida Department of Agriculture and Consumer Service Division of Plant Industry are mass rearing and releasing P. ichini in Florida. Over 4 million have been released on the landscape and populations are persisting and spreading into new areas (Wheeler et al., 2022; C. Minteer, personal communication). No non-target impacts have been seen in the field and testing of host specificity in open field situations confirm results of host-range tests that were conducted in the lab before release (Bowers et al., 2022).
Brazilian Peppertree Sawfly, Heteroperreyia hubrichi
Heteroperreyia hubrichi Malaise (Hymenoptera: Pergidae) is a primitive nonstinging wasp native to northern Argentina and southeastern Brazil. The biology, ecology, and host range of the sawfly H. hubrichi were investigated in Brazil, Florida, and Hawaii (Vitorino et al. 2000, Medal et al. 1999, Hight et al. 2002, Cuda et al. 2005).

Credit: J. C. Medal, University of Florida.
Adult females mate and oviposit in young woody branches that are adjacent to the more tender terminal shoots. This behavior enables the sawfly to avoid the toxic resin common in the Brazilian peppertree's terminal growth. The female uses her saw-like ovipositor to cut the stem tissue and insert her eggs between the thin bark and the phloem, the vascular tissue that transports nutrients from the leaves (Figure 3). The eggs are elliptical in shape and are deposited side by side in long rows of variable length and number. Females exhibit maternal behavior by guarding the egg masses during the incubation period but die as soon as the first larvae hatch.
The larva of H. hubrichi is the damaging stage. Developing larvae are voracious leaf feeders (Figure 3) and can cause complete defoliation of Brazilian peppertrees depending on the size of the plant and quantity of larvae present. This type of feeding damage could severely injure or kill young plants and prevent older plants from reproducing, which would reduce the competitive advantage that Brazilian peppertree currently holds over native vegetation. In Brazil, it is not uncommon to find Brazilian peppertree shrubs and more rarely trees completely defoliated by the sawfly. Larvae are gregarious in the early instars and feed in groups on tender leaves, mainly on new shoots. When the larvae reach the third instar, they disperse over the plant and attack leaves of all age classes.
The TAG recommended field release of the defoliating sawfly H. hubrichi in Florida in 1997. However, a release permit was not issued by APHIS PPQ because of concerns raised about toxins present in the larvae, which could have a negative impact on native fauna if the larvae were ingested.
Brazilian Peppertree Leaflet Roller, Episimus unguiculus
Episimus unguiculus Clarke, previously known as E. utilis Zimmerman (Lepidoptera: Tortricidae), was introduced into Hawaii for classical biological control of Brazilian peppertree in the 1950s (Krauss 1963). Martin et al. (2004) investigated the biology of E. unguiculus in the process of establishing a laboratory colony for conducting host range tests in an approved Florida containment laboratory. Adults (Figure 4) are small, grayish brown moths with distinctive markings on the forewings. When at rest, the adults are cryptically colored, resembling either tree bark or bird droppings. Sexes can be readily separated without magnification by examining the wing pattern. Average life span for the adult moths is 8 to 9 days, and development from egg to adult stage occurs in about 42 days.

Credit: Photo of larva by M. Fukada, Hawaii Department of Agriculture (Used with permission).
Females can deposit up to 172 eggs during their lives. Eggs are usually deposited singly but occasionally in groups of up to six eggs on the upper and lower surfaces of Brazilian peppertree leaflets. The eggs, which are glued to the leaflet, are compressed, ovoid, and light green in color with a smooth chorion when first deposited but darken as they develop. The thin, scalelike shape and transparency of freshly deposited eggs probably afford them some protection from predation and possibly parasitism.
The caterpillar (or larval stage) of E. unguiculus (Figure 4) attacks the foliage of Brazilian peppertree. Feeding habits of the larvae vary depending upon their age. Newly hatched larvae and early instars feed by scraping the surface of the leaflets. Early instars are leaflet tiers, and normally feed between young and expanding leaflets that have been tied together with silk. The first to third instars typically web together two or more adjacent leaflets flat against each other. Older larvae bind single leaflets into the characteristic cylindrical roll that is usually associated with E. unguiculus in nature. A cohort of approximately 35 larvae is capable of completely defoliating a 0.5 m tall Brazilian peppertree potted plant in less than three weeks (Martin et al. 2004). The results of a recent study by Manrique et al. (2009) showed that high levels of defoliation by E. unguiculus significantly reduced the number of leaflets, plant height, foliar biomass, relative growth rate (RGR), and shoot:root ratio of potted Brazilian peppertrees. Moreover, plants were not able to recover from the effects of the herbivory after 2 months.
In Hawaii, where it was released in the 1950s, E. unguiculus is widely distributed on Brazilian peppertree, but the insect apparently is not sufficiently abundant to severely damage the plant (Yoshioka and Markin 1991; J.P. Cuda 2002, personal observation). The ineffectiveness of E. unguiculus as a biological control agent in the Hawaiian Islands may be due in part to biotic mortality factors unique to that environment. For example, two wasps that were introduced into Hawaii for classical biological control of the sugar cane leafroller Hedylepta (=Omiodes) accepta (Butler) were discovered attacking E. unguiculus soon after it was released against Brazilian peppertree (Krauss 1963).
Although satisfactory biological control of Brazilian peppertree by E. unguiculus was not achieved in the Hawaiian archipelago, this failure should not preclude the introduction of the insect into Florida. Episimus unguiculus could be a more effective biological control agent of Brazilian peppertree in Florida because it would be introduced into a new environment where biotic mortality from introduced and native parasitoids and predators may be less severe compared to Hawaii. Host range testing was completed, and a release petition was submitted to the TAG in September 2009, later revised and resubmitted in September 2012 (Cuda et al. 2019). However, TAG did not recommend field release in Florida because of the risk of apparent competition with native Episimus spp., and also the potential for the insect to attack cultivated pistachio in the unlikely event that E. unguiculus emigrated to southern California and Arizona. The risk to nontarget insects and plants from E. unguiculus could be field tested using the F1 Sterile Insect Technique (Cuda 2016). Production of F1 sterile progeny would be obtained by exposing the parental generation to low doses of gamma radiation (Moeri et al. 2009). This would facilitate observations on survival and impact of the F1 larvae on nontarget plants under actual field conditions with no risk of permanent establishment, because the adults produced from the F1 larvae are completely sterile.
Brazilian Peppertree Stem Boring Weevil, Apocnemidophorus pipitzi
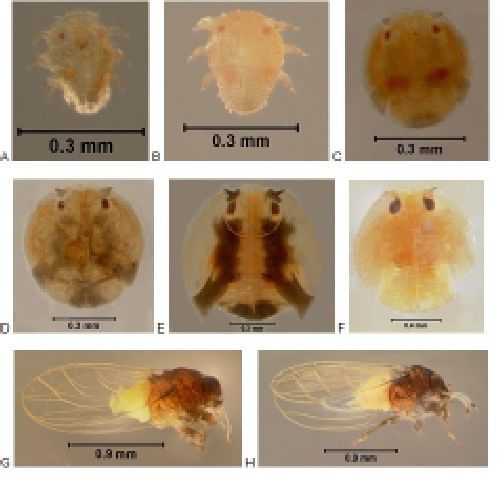
Surveys conducted in northern Argentina (McKay et al. 2009), and southeastern Paraguay revealed the presence of several new natural enemies of Brazilian peppertree. Among these were two species of stem boring weevils belonging to the genus Apocnemidophorus Hustache. The weevils resemble bird droppings and immediately fall off the plants when they are disturbed. Adults of Apocnemidophorus pipitzi (Faust) were collected in southeastern Paraguay and imported into a Florida containment laboratory in April 2006. According to Wibmer and O'Brien (1986), A. pipitzi is native to Argentina, Brazil, and Uruguay. An examination of weevil specimens deposited in the entomology museum of the Federal University of Parana in Curitiba, Brazil, revealed that collections of A. pipitzi have been made in the states of Parana, Santa Catarina, and Minas Gerais. The occurrence of A. pipitzi in Paraguay is a new country record for this species.

Credit: Lyle Buss, University of Florida.
The life stages of A. pipitzi are shown in Figure 5. The adults are long-lived, surviving in the laboratory for almost 2 months (Cuda et al. 2016). Females chew small holes (0.5 mm) into the stems and then insert their ovipositors into the cavities. After depositing an egg (or multiple eggs), they seal the cavities with a frass plug that initially is bright green in color but eventually turns brown. The entire process lasts approximately 45 minutes. In the laboratory, a new generation of weevils emerged from cut stems of Brazilian peppertree in 3 to 4 months (Cuda et al. 2016).
Both the adults and larvae of A. pipitzi are capable of damaging Brazilian peppertree (Cuda et al. 2016). Adults feed preferentially on the tender, subterminal leaflets of the plant. The characteristic feeding scars produced by the adults generally do not completely perforate the leaflets, causing a thin, translucent layer of plant tissue or "window" to remain. However, leaflets that sustain heavy feeding damage eventually abscise. The larvae (Figure 5) tunnel extensively just under the bark of BP (in the vicinity of the cambium) until they reach the pupal stage. This type of feeding damage can weaken the plant because it interferes with the normal transport of water and nutrients in the xylem and phloem tissues. A petition for release of A. pipitzi was submitted in October 2012 (Cuda et al., 2021). However, the TAG did not recommend this species for release in Florida and requested additional host range tests. Additional host range tests were completed and found that A. pipitzi has the ability to complete development on Brazilian peppertree, Hardee peppertree (Schinus polygama (Cav.) Cabrera), and Chinese pistache (Pistacia chinensis Bunge) and pistachio (Pistacia vera L.) (Cuda et al., 2021).
Brazilian Peppertree Leaf Gallers, Calophya spp.
Three other promising insects that have biological control potential are the leaflet galling psyllids Calophya terebinthifolii Burckhardt and Basset (Figure 6) (Vitorino et al. 2011, Christ et al. 2013) and the congeners Calophya latiforceps Burckhardt and Calophya lutea Burckhardt (Hemiptera: Calophyidae) (Burckhardt et al. 2011, 2018, Diaz et al. 2014a). These three Calophya spp., which are native to Brazil and Paraguay, create open pit galls on the leaflets of Brazilian peppertree as they complete their development to the adult stage. Results of field and laboratory studies in Brazil indicate that these insects are Schinus specialists (Vitorino et al. 2011, Diaz et al. 2014b. Burckkardt et al. 2018; Prade et al., 2021). Females live between 9 to 11 days and deposit 55 to 86 eggs. A new generation is produced in about 44 days. At high densities, these psyllids cause leaf abscission and can stunt the growth of Brazilian peppertree (Prade et al. 2015). A petition to release C. latiforceps from quarantine was submitted to the TAG in April 2015, TAG recommended field release in April 2016, and a release permit was issued in June 2019. No releases of this species have been made as of 2022 due to the loss of the colony during the COVID-19 pandemic and the inability to return to Brazil to collect this insect from the field.
Credit: Lindsey R. Christ, University of Florida.
References
Bowers, K., S. D. Hight, G. S. Wheeler, C. R. Minteer. 2022. Ecological host range of Pseudophilothrips ichini (Thysanoptera: Phlaeothripidae), a biological control agent of Brazilian peppertree, Schinus terebinthifolia. Biological Control, 172: 104976.
Burckhardt, D., J. P. Cuda, V. Manrique, R. Diaz, W. A. Overholt, D. A. Williams, L. R. Christ, and M. D. Vitorino. 2011. Calophya latiforceps, a new species of jumping plant lice (Hemiptera: Calophyidae) associated with Schinus terebinthifolius (Anacardiaceae) in Brazil. Florida Entomol. 94(3): 489–499. https://doi.org/10.1653/024.094.0313
Burckhardt, D., J. P. Cuda, R. Diaz, W. Overholt, P. Prade, D. L. de Queiroz, M. D. Vitorino, and G. S. Wheeler. 2018. "Taxonomy of Calophya (Hemiptera: Calophyidae) species associated with Schinus terebinthifolia (Anacardiaceae)." Florida Entomologist 101: 178–188.
Christ, L.R., J.P. Cuda, W.A. Overholt, M.D. Vitorino, and A. Mukherjee. 2013. Biology, host preferences, and potential distribution of Calophya terebinthifolii (Hemiptera: Calophyidae), a candidate for biological control of Brazilian peppertree, Schinus terebinthifolius, in Florida. Florida Entomol. 96: 137–147. https://doi.org/10.1653/024.096.0118
Cuda, J.P. 2016. Novel approaches for reversible field releases of candidate weed biological control agents: Putting the genie back into the bottle, Chapter 7, pp. 137-152. In: J.F. Shroder and R. Sivanpillai (eds.), Biological and Environmental Hazards, Risks and Disasters. Elsevier, Inc., Amsterdam. https://doi.org/10.1016/B978-0-12-394847-2.00010-3
Cuda J.P., A.P. Ferriter, V. Manrique, and J.C. Medal (eds.). 2006. Florida's Brazilian peppertree management plan, 2nd edition: Recommendations from the Brazilian peppertree Task Force, Florida Exotic Pest Plant Council, April 2006. Available at https://ipm.ifas.ufl.edu/pdfs/BPmanagPlan.pdf. Accessed August 4, 2022.
Cuda, J.P., D.H. Habeck, S.D. Hight, J.C. Medal, and J.H. Pedrosa-Macedo. 2004. Brazilian Peppertree, Schinus terebinthfolius: Sumac Family-Anacardiaceae, pp. 439-441. In E. Coombs, J. Clark, G. Piper, and A. Cofrancesco (eds.), Biological Control of Invasive Plants in the United States. Oregon State University Press, Corvallis, OR.
Cuda, J.P., J.C. Medal, J.H. Pedrosa-Macedo, and D.H. Habeck. 2002. Request for field release of a nonindigenous thrips Pseudophilothrips ichini (Thysanoptera: Phlaeothripidae) for classical biological control of Brazilian peppertree, Schinus terebinthifolius (Anacardiaceae), in Florida (Submitted to IFAS and TAG October 2002). 53 pp.
Cuda, J.P., J.C. Medal, J.L. Gillmore, D.H. Habeck, and J.H. Pedrosa-Macedo. 2009. Fundamental host range of Pseudophilothrips ichii sensu lato (Thysanoptera: Phlaeothripidae), a Anacardiaceae) in the USA. Environ. Entomol. 38: 1642–1652. https://doi.org/10.1603/022.038.0617
Cuda, J.P., J.C. Medal, M.D. Vitorino, and D.H. Habeck. 2005. Supplementary host specificity testing of the sawfly Heteroperreyia hubrichi, a candidate for classical biological control of Brazilian peppertree, Schinus terebinthifolius, in the USA. BioControl 50: 195–201. https://doi.org/10.1007/s10526-004-0459-3
Cuda, J.P., J.L. Gillmore, A.O. Mitchell, J. Bricker, R.A. Watson, B.R. Garcete-Barrett, and A. Mukherjee. 2016. Laboratory biology and impact of a stem boring weevil Apocnemidophorus pipitzi (Faust) (Coleoptera: Curculionidae) on Schinus terebinthifolia. Biocontrol Science and Technology 26: 1249-1266. https://doi.org/10.1080/09583157.2016.1193844
Cuda, J.P., J. L. Gillmore, P. Conant, J. C. Medal, and J. H. Pedrosa-Macedo. 2019. Risk Assessment of Episimus unguiculus (Lepidoptera: Tortricidae), a Biological Control Agent of Schinus terebinthifolia (Sapindales: Anacardiaceae) in Hawaii, USA. Biocontrol Science and Technology 29(4): 365-387. DOI: 10.1080/09583157.2018.1562041
Cuda, J.P., J. L. Gillmore, B. R. Garcete-Barrett, N. Benda, S. Sharma. 2021. Is the stem boring weevil Apocnemidophorus pipitzi (Coleoptera: Curculionididae) host specific to Schinus terebinthifolia (Sapindales: Anacardiaceae)?. Biocontrol Science and Technology, 31: 1128-1148.
Diaz R, Moscoso D, Manrique V, Williams D, Overholt WA. 2014a. Native range density, host utilization and life history of Calophya latiforceps (Hemiptera: Calophyidae): an herbivore of Brazilian peppertree (Schinus terebinthifolia). Biocontrol Sci & Tech. 24: 536–553. https://doi.org/10.1080/09583157.2013.878686
Diaz R, Manrique V, Munyaneza JE, Sengoda VG, Adkins S, Hendricks K, Roberts PD, Overholt WA. 2014b. Host specificity testing and examination for plant pathogens reveal that the gall-inducing psyllid Calophya latiforceps is safe to release for biological control of Brazilian peppertree. Entomol Experimentalis et Applicata 1-14 https://doi.org/10.1111/eea.12249
Furmann, L.E., J.H. Pedrosa-Macedo, J.P. Cuda, and M.D. Vitorino. 2005. Efeito da liberação no campo de Pseudophilothiprs ichini no desenvolvimento de Schinus terebinthifolius. Floresta 35: 241–245. https://doi.org/10.5380/rf.v35i2.4610
Garcia, C.A. 1977. Biologia e aspectos da ecologia e do comportamento defensiva comparada de Liothrips ichini Hood 1949 (Thysanoptera Tubulifera). M.S. Thesis, Universidade Federal do Parana, Curitiba, Parana, Brazil. 75 p.
Habeck, D.H., F.D. Bennett, and J.K. Balciunas. 1994. Biological control of terrestrial and wetland weeds, pp. 523-547. In D. Rosen, F.D. Bennett, and J.L. Capinera (eds.), Pest Management in the Subtropics: Biological Control — a Florida Perspective. Intercept, Andover, United Kingdom.
Hiatt, D., K. Serbesoff-King, D. Lieurance, D.R. Gordon and S. Luke Flory. 2019. Allocation of invasive plant management expenditures for conservation: Lessons from Florida, USA. Conservation Science and Practice 1: e51. 10 pp. https://doi.org/10.1111/csp2.51
Hight, S.D., J.P. Cuda, and J.C. Medal. 2002. Brazilian peppertree. Pages 311-321 In R. Van Driesche, B. Blossey, M. Hoddle, S. Lyon, and R. Reardon (eds.). Biological Control of Invasive Plants in the Eastern United States. US Forest Service, Morgantown, WV, FHTET-2002-04.
Krauss, N.L.H. 1963. Biological control investigations on Christmas berry (Schinus terebinthifolius) and emex (Emex spp.). Proc. Hawaiian Entomol. Soc. 18: 281-288.
Manrique V, Cuda J.P., Overholt W.A., Williams D.A., and G. S. Wheeler. 2008. Effect of host-plant genotypes on the performance of three candidate biological control agents of Schinus terebinthifolius in Florida. Biological Control 47: 167–171.
Manrique, V. J. P. Cuda, and W. A. Overholt. 2009. Effect of herbivory on growth and biomass allocation of the invasive Brazilian peppertree (Sapindales: Anacardiaceae) seedlings in the laboratory. BioControl Sci. & Tech. 19: 657–667.
Manrique, V., J.P. Cuda, and W.A. Overholt. 2013. Brazilian peppertree: a poster child for invasive plants in Florida landscapes. Journal of Florida Studies, 1, 1–14. Retrieved from https://www.journaloffloridastudies.org/0102peppertree.html
Martin, C.G., J.P. Cuda, K.D. Awadzi, J.C. Medal, D.H. Habeck and J.H. Pedrosa-Macedo. 2004. Biology and laboratory rearing of Episimus utilis (Lepidoptera: Tortricidae), a candidate for classical biological control of Brazilian peppertree, Schinus terebinthifolius (Anacardiaceae), in Florida. Environ. Entomol. 33 1351–1361. https://doi.org/10.1603/0046-225X-33.5.1351
McKay, F., M. Oleiro, G.C. Walsh, D. Gandolfo, G.S. Wheeler, and J.P. Cuda. 2009. Natural enemies of Brazilian peppertree (Schinus terebinthifolius: Anacardiaceae) from Argentina: their possible use for biological control in the USA. Florida Entomol. 92: 292–303. https://doi.org/10.1653/024.092.0213
Medal, J.C., M.D. Vitorino, D.H. Habeck, J.L. Gillmore, J.H. Pedrosa, and L.D. De Sousa. 1999. Host specificity of Heteroperreyia hubrichi Malaise (Hymenoptera: Pergidae), a potential biological control agent of Brazilian Peppertree (Schinus terebinthifolius Raddi). Biological Control 14: 60–65. https://doi.org/10.1006/bcon.1998.0670
Moeri, O.E. J. P. Cuda, W.A. Overholt, S. Bloem, and J. E. Carpenter. 2009. F1 Sterile Insect Technique: a Novel Approach for Risk Assessment of Episimus unguiculus (Lepidoptera: Tortricidae), a Candidate Biological Control Agent of Schinus terebinthifolius in the Continental USA. BioControl Science and Technology 19, Supplement 1: 303–315. https://doi.org/10.1080/09583150902741932
Mound, L.A., and R. Marullo. 1996. The thrips of Central and South America: an introduction (Insecta: Thysanoptera). Memoirs Entomol. International 6: 1-487.
Mound, L.A. G.S. Wheeler, and D.A. Williams. 2010. Resolving cryptic species with morphology and DNA; thrips as a potential biocontrol agent of Brazilian peppertree, with a new species and overview of Pseudophilothrips (Thysanoptera). Zootaxa 2432: 59–68.
Prade, P, R. Diaz, M. D. Vitorino, J. P. Cuda, P. Kumar, B. Gruber, W.A. Overholt. 2015. Galls induced by Calophya latiforceps Burckhardt (Hemiptera: Calophyidae) reduce leaf performance and growth of Brazilian peppertree. Biocontrol Science and Technology 26: 23-34. https://doi.org/10.1080/09583157.2015.1072131
Prade, P., C. R. Minteer, S. A. Gezan, V.C. Arguijo, K. Bowers, J. P. Cuda, W. A. Overholt. 2021. Host specificity and non-target longevity of Calophya lutea and Calophya terebinthifolii, two potential biological control agents of Brazilian peppertree in Florida, USA. BioControl, 66: 281-294.
Rodgers, L., T. Pernas, and S D. Hill, 2014. Mapping invasive plant distributions in the Florida Everglades using the digital aerial sketch mapping technique. Invasive Plant Science and Management, 7, 360-374. https://doi.org/10.1614/IPSM-D-12-00092.1
Treadwell, L.W. and J.P. Cuda., J.P. 2007. Effects of defoliation on growth and reproduction of Brazilian peppertree (Schinus terebinthifolius). Weed Science 55: 137-142. https://doi.org/10.1614/WS-06-086.1
Vitorino, M.D., J.H. Pedrosa-Macedo, and J.P. Cuda. 2000. Biology and specificity tests of the sawfly – Heteroperreyia hubrichi Malaise, 1955 (Hymenoptera: Pergidae) a potential biological control agent for Brazilian peppertree- Schinus terebinthifolius Raddi (Anacardiaceae). In: N.R. Spencer (ed.), Proceedings of the X International Symposium on Biological Control of Weeds, 4-14 July 1999, Montana State University, Bozeman, Montana, US. USDA, ARS, Sidney, Montana and Montana State University, Bozeman, Montana. pp. 645–650.
Vitorino, M.D., L.R. Christ, G. Barbieri, J.P. Cuda and J.C. Medal. 2011. Calophya terebinthifolii (Hemiptera: Psyllidae), a candidate for biological control of Schinus terebinthifolius (Anacardiaceae): Preliminary host range, dispersal, and impact studies. FL Entomol. 94: 694–695. https://doi.org/10.1653/024.094.0337
Wheeler, G.S., N. Silverson, K. Dyer, F. Mc Kay. 2016. Brazilian collections and laboratory biology of the thrips Pseudophilothrips ichini (Thysanoptera: Phlaeothripidae): a potential biological control agent of the invasive weed Brazilian peppertree (Sapindales: Anacardiaceae). FL Entomol. 99: 6–11. https://doi.org/10.1653/024.099.0103
Wheeler. G.S., V. Manrique, W.A. Overholt, F. McKay and K. Dyer. 2017. Quarantine host range testing of Pseudophilothrips ichini, a potential biological control agent of Brazilian peppertree, Schinus terebinthifolia, in North America and Hawaii. Entomol. Exp. Appl. 162: 204-217.
Wibmer, G.J. and C.S. OBrien. 1986. Annotated checklist of the weevils (Curculionidae sensu lato) of South America (Coleoptera: Curculionoidea). Mem. Amer. Entomol. Institute 39. American Entomological Institute, Gainesville, FL.
Williams, J. R. 1954. The biological control of weeds. In: Report of the Sixth Commonwealth Entomological Congress, London, UK, pp. 95–98.
Yoshioka, E.R., and G.P. Markin. 1991. Efforts of biological control of Christmas berry Schinus terebinthifolius in Hawaii. pp. 377-385. In T. Center et al. (eds.), Proceedings of the symposium of exotic pest plants, 2–4 November 1988. Miami, FL.

